Search this article
Dr. Elemans et al. replicate bird song using pressure in a closed system
How do birds sing? Much like you and I do, it turns out.
Research by Dr. Coen Elemans et al, Universal mechanisms of sound production and control in birds and mammals, published in Nature Communications Nov. 27, 2015, found that birds and mammals use the same physical mechanism of sound production – the myoelastic-aerodynamic (MEAD) mechanism.
Crucial to this research was the ability to replicate tissue oscillation in the syrinx, or voice box – a bird’s correlate to our larynx – by finely controlling the pressures in a closed system below and outside this organ. To gain this control, Dr. Elemans used Alicat’s dual-valve pressure controllers for their high degree of accuracy and control within closed volumes.
Pressure control animates MEAD oscillations
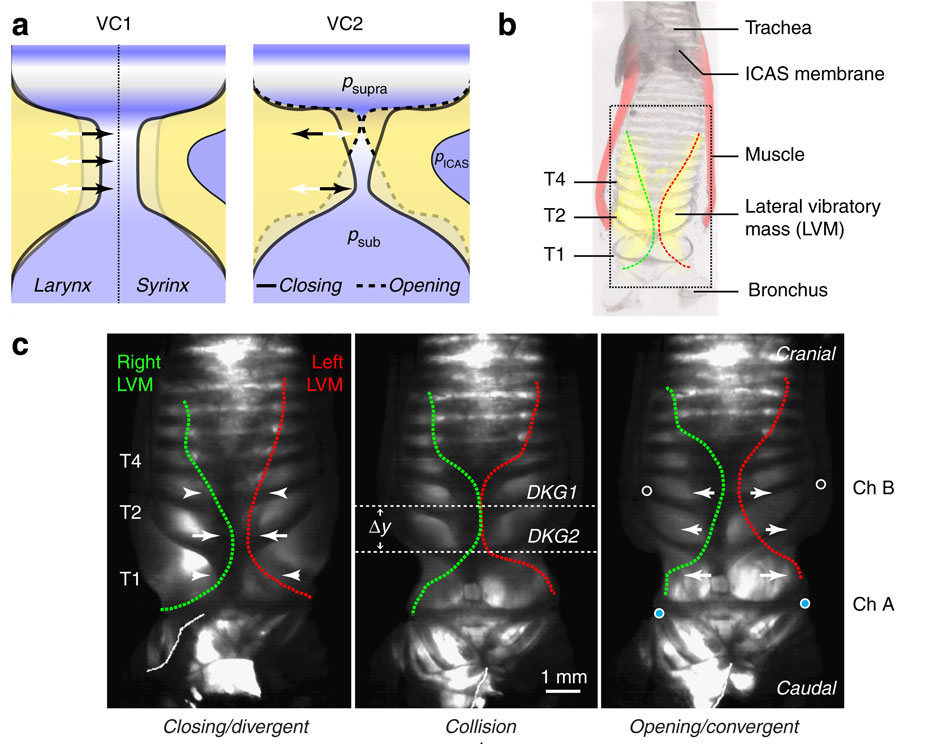
A portion of Figure 1 from the team’s article in Nature, showing MEAD oscillations. Click to see the full image.
Animal vocal sounds are composed of discrete pulses of air generated by vocal tissue oscillations at a frequency of several hundred times per second. The MEAD mechanism describes how these oscillations are sustained without the need for active muscle vibrations at the same frequency, which would surely tire out any animal (the fastest known muscles cannot contract faster than 250 Hz.)
In MEAD theory, air pressure builds below the closed vocal tissue until there is enough pressure to force the tissue to open. The tissue opens and closes asymmetrically, and the passing air is chopped up by its oscillations, making sound. The frequency of the oscillations determines the pitch of the sound, and the oscillation frequency is determined by the flow rate through the voice box and the muscular tension of its tissue.
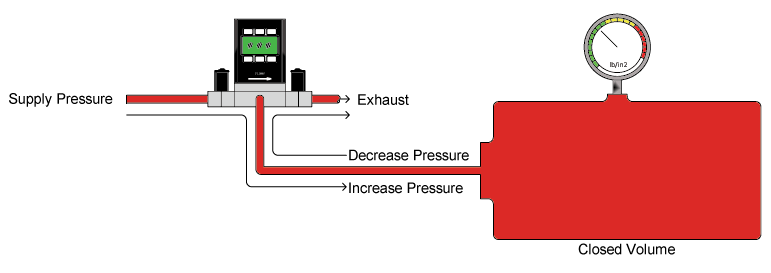
To determine whether MEAD functioned in birds as it had been demonstrated to operate in mammals, Dr. Elemans’ team had to replicate pressure-induced airflow through the syrinx.
The team wanted to precisely control pressure in the bronchial airways below the vocal tissue to test the presence of MEAD oscillations. Because the remainder of the bird’s respiratory system was effectively dead-ended, Alicat’s dual-valve pressure controller was the perfect solution. The two valves, one at the inlet and the other at the oulet, allow the device to maintain exact pressures in a closed system by adding or removing air from the volume as needed.

Differential pressure controller for closed volumes with remote display. Note the dual pressure sense ports on the front.
The research team also needed to control up to 3 kPA (0.4 PSIG), a low amount of positive pressure, to prevent damage to the biological structures they were studying. For this, Alicat specified the use of a differential pressure sensor within the pressure controller. One of the remote sense ports was plumbed to the bronchial airways of the bird, and the other was left open to atmosphere.
This allowed the bronchial pressure to be constantly referenced to the local atmospheric pressure, no matter how slightly it changed throughout the experiment. Integrated analog and digital control signals made it easy for the team to subject the syrinx to bronchial pressure ramps.
Conclusions: Pressure control spaces and redundancy
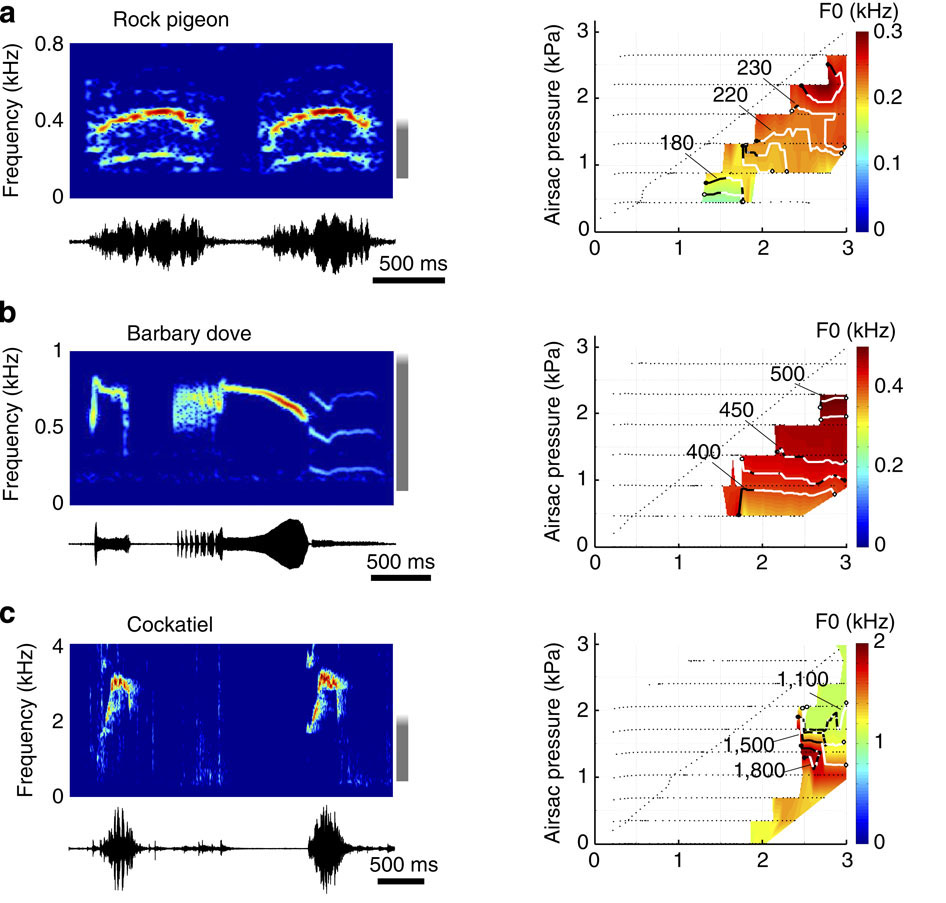
A portion of Figure 6 from the team’s article in Nature, showing redundant pressure control spaces. Click to see the full image.
In addition to proving the operation of MEAD within birds, Dr. Elemens also wanted to determine whether bird vocalizations resulted from unique muscle commands or a redundant control space. To test this, the team varied the interclavicular air sac (ICAS) pressure that surrounds the syrinx, while simultaneously subjecting the syrinx to bronchial pressure ramps and varying degrees of muscle stimuli. A second PCD was employed to vary the simulated ICAS pressure within the same 1-3 kPa range as the bronchial volume.
The team found that multiple combinations of pressures within the two pressure zones, as well as muscular stimuli, were able to yield the same fundamental frequency, a redundancy feature that is common to MEAD.
Significantly, Dr. Elemens’ team also found that the aerodynamic engine that sustained the vocal tissue oscillations was not generated by “the mass inertance of air column in vocal tract, but by the tissue-wave-induced intraglottal pressure changes” (page 6 of their study). Alternating low and high pressures in order to maintain flow through the vocal tissue was not caused by the mass of air moving up the bird’s throat. Instead, the wave motion of the tissue edges generated the necessary pressure changes within the syrinx. This created a region of low pressure in the closed system until the tissue opened again.